Bewerkt door Dr. Giovanni Chetta
Van biochemie naar biomechanica

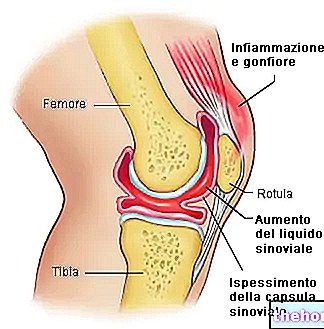
Extracellulaire matrix (MEC)
Een beschrijving, zij het van het weinige dat we vandaag weten, van de ECM (extracellulaire matrix) is essentieel om het belang van veranderingen in de wervelkolom en houding voor de gezondheid beter te begrijpen.
Elke cel, zoals elk meercellig levend organisme, moet zijn omgeving "voelen" en ermee omgaan om zijn vitale functies uit te voeren en te overleven. In een meercellig organisme moeten de cellen de verschillende gedragingen coördineren zoals in een gemeenschap van mensen.

ECM wordt over het algemeen beschreven als zijnde samengesteld uit verschillende grote klassen van biomoleculen:
- Structurele eiwitten (collageen en elastine)
- Gespecialiseerde eiwitten (fibrilline, fibronectine, laminine enz.)
- Proteoglycanen (aggrecanen, syndecanen) en glusaminoglycanen (hyaluronanen, chondroïtinesulfaten, heparansulfaten, enz.)
Onder de structurele eiwitten, i collageen vormen de meest vertegenwoordigde familie van glycoproteïnen in het dierenrijk. Het zijn de meest aanwezige eiwitten in de ECM (maar niet de belangrijkste) en zijn de fundamentele bestanddelen van het eigenlijke bindweefsel (kraakbeen, bot, fascia, pezen, ligamenten).
Collagenen worden meestal gesynthetiseerd door fibroblasten, maar epitheelcellen kunnen ze ook synthetiseren.
De collageenvezels staan continu in wisselwerking met een enorme hoeveelheid andere moleculen van de ECM, die een biologisch continuüm vormen dat fundamenteel is voor het leven van de cel. De geassocieerde collagenen in fibrillen spelen een overheersende rol bij de vorming en het onderhoud van structuren die in staat zijn om spanningskrachten te weerstaan, die bijna inelastisch Collageen wordt geproduceerd en opnieuw gemetaboliseerd als functie van de mechanische belasting en zijn visco-elastische eigenschappen hebben een grote invloed op de houding van de mens.
Collageenvezels hebben dankzij hun PG / GAG-coating (proteoglycanen / glucosaminoglycanen) biosensor- en biogeleidende eigenschappen. We weten in feite dat elke mechanische kracht die een structurele vervorming kan veroorzaken, de intermoleculaire bindingen onder druk zet en een lichte elektrische flux produceert, dat wil zeggen de piëzo-elektrische stroom (Athenstaedt, 1969). Daarom bezit het driedimensionale en alomtegenwoordige collageennetwerk ook het bijzondere kenmerk van het geleiden van bio-elektrische signalen in de drie dimensies van de ruimte, gebaseerd op de relatieve rangschikking tussen collageenfibrillen en cellen, in de afferente richting (van de ECM naar de cellen) of vice versa. omgekeerd efferent.
Dit alles vertegenwoordigt een real-time MEC-celcommunicatiesysteem en dergelijke elektromagnetische biosignalen kunnen leiden tot belangrijke biochemische veranderingen, bijvoorbeeld in "botosteoclasten kunnen" piëzo-elektrisch geladen bot niet verteren (Oschman, 2000).
Ten slotte moet worden benadrukt dat de cel, niet verrassend, continu en met een aanzienlijk energieverbruik (ongeveer 70%) materiaal produceert dat noodzakelijkerwijs moet worden verdreven door de exclusieve opslag van protocollagen (biologische voorloper van collageen) in specifieke blaasjes ( Albergati, 2004).
De overgrote meerderheid van gewervelde weefsels vereist de gelijktijdige aanwezigheid van twee vitale kenmerken: sterkte en elasticiteit. Een echt netwerk van elastische vezels, die zich in de ECM van deze weefsels bevindt, maakt het mogelijk om na sterke tracties terug te keren naar de oorspronkelijke omstandigheden. De elastische vezels zijn in staat om de rekbaarheid van een orgaan of een deel ervan ten minste vijf keer te vergroten. Lange, niet-elastische collageenvezels worden afgewisseld tussen de elastische vezels met de precieze taak om een "overmatige vervorming door tractie van de weefsels" te beperken. vormt meer dan 50% van het totale droge gewicht van de aorta), in de ligamenten, in de longen en in de huid Gladde spiercellen en fibroblasten zijn de belangrijkste producenten van zijn voorloper, tropoelastine.
ECM bevat een groot (en nog steeds niet goed gedefinieerd) aantal gespecialiseerde niet-collageeneiwitten die typisch specifieke bindingsplaatsen hebben voor andere ECM-moleculen en celoppervlakreceptoren. Op deze manier fungeert elk afzonderlijk onderdeel van deze eiwitten als een "versterker" van contacten, zowel tussen vergelijkbare als verschillende moleculen, waardoor een oneindig biochemisch netwerk ontstaat dat in staat is om zelfs op een afstand miljoenen en miljoenen biochemische informatie te genereren, moduleren, variëren en verspreiden. (en energie).
Een belangrijk "gespecialiseerd eiwit van de extracellulaire matrix is de" fibronectine, hoogmoleculair glycoproteïne gevonden in alle gewervelde dieren. Fibronectine blijkt celgroei, intercellulaire adhesie en met ECM celmigratie op verschillende manieren te kunnen beïnvloeden (de cel kan tot 5 cm per dag bewegen - Albergati, 2004) etc. De bekendste isovorm, type III, bindt aan integrines . Deze laatste zijn een familie van transmembraaneiwitten die fungeren als mechanoreceptoren: ze transduceren, selectief en op een moduleerbare manier, mechanische tracties en stoten van de ECM in de cel en vice versa, en induceren een reeks reacties in het cytoplasma waarbij het cytoskelet en andere eiwitten die ze celadhesie, groei en migratie reguleren (Hynes, 2002).
Glucosaminoglycanen (GAGS) en proteoglycanen (PG's) vormen een sterk gehydrateerde gelachtige substantie die wordt gedefinieerd in de bindweefsels, waarin de fibrillaire eiwitten zijn ondergebracht en overlappen. Deze vorm van polysacharidegel is enerzijds in staat om de ECM in staat te stellen aanzienlijke drukkrachten te weerstaan en anderzijds om een snelle, constante en selectieve diffusie van voedingsstoffen, metabolieten en hormonen tussen bloed en weefsels mogelijk te maken.
De polysacharideketens van glucosaminoglycanen zijn volumetrisch te rigide om te vouwen in de compacte bolvormige structuren die typisch zijn voor polypeptideketens, bovendien zijn ze zeer hydrofiel.Om deze redenen (en waarschijnlijk ook voor anderen die ons onbekend zijn) hebben de GAG's de neiging om extreem conformaties aan te nemen. een groot volume in verhouding tot hun massa innemen en aldus aanzienlijke hoeveelheden gel vormen, zelfs bij lage concentraties.De grote hoeveelheid negatieve ladingen (de GAG's vertegenwoordigen de meest talrijke anionische cellen, meestal gesulfateerd, geproduceerd door dierlijke cellen) trekt talrijke kationen aan; onder deze wordt een overheersende rol gespeeld door Na + dat de hele osmotische capaciteit geeft en een enorme hoeveelheid water in de ECM vasthoudt. Op deze manier worden zwellingen (turgors) gegenereerd die de ECM in staat stellen om zelfs belangrijke compressiekrachten te weerstaan (hierdoor kan bijvoorbeeld het kraakbeen van de heup onder fysiologische omstandigheden een druk van enkele honderden atmosfeer perfect weerstaan).
In het bindweefsel vertegenwoordigen de GAG's minder dan 10-12% van het totale gewicht, maar dankzij hun kenmerken vullen ze veel van de extracellulaire ruimten en vormen ze poriën van gehydrateerde gel van verschillende groottes en dichtheden van elektrische ladingen, waardoor ze als selectief werken sleutelpunten of "servers" waarmee het verkeer van moleculen en cellen binnen de MEC wordt geregeld, op basis van hun grootte, gewicht en elektrische lading.
Hyaluronzuur (hyaluronzuur, hyaluronaat) vertegenwoordigt misschien wel de eenvoudigste van de GAG's Experimentele en moleculair-biologische gegevens bevestigen dat het een fundamentele rol speelt op het niveau van botten en gewrichten wat betreft weerstand tegen aanzienlijke druk opvulling van ruimtes in de ECM tijdens de embryonale ontwikkeling : het creëert lege ruimtes tussen de cellen waarin ze in latere stadia zullen migreren (Albergati, 2004).
Niet alle PG's worden uitgescheiden door ECM, sommige zijn integrale componenten van plasmamembranen (Alberts, 2002).
De extracellulaire matrix kan daarom worden beschouwd als een zeer complex netwerk waarin eiwitten, PGS en GAG's ontelbare functies bieden, waaronder die van structurele ondersteuning en regulatie van elk weefsel en organische activiteit. Globale cellulaire homeostase moet worden beschouwd als een complex van mechanismen die binnen de cel of buiten de ECM kunnen ontstaan en zich kunnen ontwikkelen; in het laatste geval kan de cel het tussenliggende of uiteindelijke doelwit vertegenwoordigen. De extracellulaire componenten vertegenwoordigen niet alleen fysieke ondersteuningsstructuren voor de cellulaire steiger, maar fungeren ook als echte plaatsen voor de initiatie, ontwikkeling en beëindiging van vitale processen met betrekking tot zowel de endocellulaire omgeving als organen en systemen. We worden geconfronteerd met een oneindig biochemisch netwerk dat in staat is miljoenen en miljoenen informatie te genereren, te moduleren, te variëren en te verspreiden, zelfs op afstand.
Elke cel van het lichaam staat voortdurend in wisselwerking met de ECM, zowel onder mechanische als chemische en energetische aspecten, met "dramatische" effecten op de statische en dynamische architectuur van de weefsels. Volgens P. A. Bacci vertegenwoordigt de interstitiële matrix echt de moeder van vitale reacties, de plaats waar allereerst de uitwisselingen tussen materie en energie plaatsvinden. Alle weefsels zijn met elkaar verbonden en functioneel geïntegreerd, niet in gesloten maar open systemen; Er vinden continue uitwisselingen tussen hen plaats, die zowel lokaal als systemisch kunnen plaatsvinden, waarbij gebruik wordt gemaakt van biochemische, biofysische en elektromagnetische berichten, dat wil zeggen met behulp van de verschillende vormen van energie.
Als F.G.Gehuisvest vertegenwoordigen de cel en de extracellulaire matrix twee schijnbaar gescheiden werelden die noodzakelijkerwijs voor de gehele duur van het leven op elk moment moeten interageren om op een correcte en synergetische manier te werken. Dit vereist een buitengewone reeks signalen, gevolgd door een even ongelooflijke reeks moleculair-biologische activiteiten.
Andere artikelen over "Extra-cellulaire matrix - Structuur en functies"
- Behandeling van scoliose
- Scoliose - Oorzaken en gevolgen
- Diagnose van scoliose
- Prognose van scoliose
- Bindweefsel en bindweefsel
- Verbindingsband - Kenmerken en functies
- Houding en tensegrity
- De beweging van de mens en het belang van stuitligging
- Belang van correcte stuitligging en occlusale steunen
- Idiopathische scoliose - Mythen om te verdrijven
- Klinisch geval van scoliose en therapeutisch protocol
- Behandelingsresultaten Klinische casus Scoliose
- Scoliose als een natuurlijke houding - Bibliografie