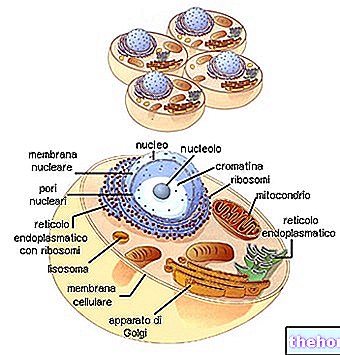
Om een overeenkomst te hebben tussen de informatie van het polynucleotide en die van het polypeptide, is er een code: de genetische code.
De algemene kenmerken van de genetische code kunnen als volgt worden opgesomd:
De genetische code bestaat uit drielingen en is verstoken van interne interpunctie (Crick & Brenner,).
Het "werd ontcijferd door het gebruik van" open cel vertaalsystemen "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Het is sterk gedegenereerd (synoniemen).
De organisatie van de codetabel is niet toevallig.
Drieling "onzin".
De genetische code is "standaard", maar niet "universeel".
Kijkend naar de tabel van de genetische code, moet worden bedacht dat deze verwijst naar de vertaling van "RNAm naar polypeptide, waarvoor de betrokken nucleotidebasen A, U, G, C zijn. De biosynthese van een polypeptideketen is de vertaling van de nucleotidesequentie in volgorde aminozuur.

Elk base-triplet van het RNAm, codon genaamd, heeft de eerste base in de linkerkolom, de tweede in de bovenste rij, de derde in de rechterkolom Laten we bijvoorbeeld tryptofaan nemen (dwz Try) en we zien dat het corresponderende codon wees, in orde, UGG. In feite omvat de eerste basis, U, de hele rij dozen bovenaan; hierin identificeert G het meest rechtse vak en de vierde regel van het vak zelf, waar we geschreven Try vinden. Evenzo kunnen we, om het tetrapeptide Leucine-Alanine-Arginine-Serína (symbolen Leu-Ala-Arg-Ser) te synthetiseren, de codons UUA-AUC-AGA-UCA in de code vinden.

Op dit punt moet echter worden opgemerkt dat alle aminozuren van ons tetrapeptide worden gecodeerd (in tegenstelling tot tryptofaan) door meer dan één codon. Het is geen toeval dat we in het zojuist gerapporteerde voorbeeld de aangegeven codons hebben gekozen, we hadden hetzelfde tripeptide kunnen coderen met een andere RNAm-sequentie, zoals CUC-GCC-CGG-UCC.
Aanvankelijk kreeg het feit dat een enkel aminozuur overeenkwam met meer dan een triplet een betekenis van willekeur, ook uitgedrukt in de keuze van de term van degeneratie van de code, die wordt gebruikt om het fenomeen synoniem te definiëren. Aan de andere kant suggereren sommige gegevens dat de beschikbaarheid van synoniemen die verwijzen naar verschillende stabiliteit van de genetische informatie helemaal niet toevallig is.Dit lijkt ook te worden bevestigd door de vondst van een andere waarde van de A + T / G + C-verhouding in de verschillende stadia van evolutie. Bij prokaryoten bijvoorbeeld, waar de behoefte aan variabiliteit niet wordt bevredigd door de regels van het mendelisme en neo-mendelisme, heeft de A + T / G + C-verhouding de neiging toe te nemen. De daaruit voortvloeiende lagere stabiliteit, in het licht van mutaties, zorgt voor een grotere mogelijkheden voor variabiliteit willekeurig van genmutatie.
Bij eukaryoten, met name in meercellige cellen, waarbij het noodzakelijk is dat de cellen van het enkele organisme allemaal hetzelfde erfelijke erfgoed behouden, heeft de A + T / G + C-verhouding in DNA de neiging af te nemen, waardoor de kans op somatische genmutaties afneemt .
Het bestaan van synonieme codons in de genetische code roept het reeds genoemde probleem op van de veelheid van anticodons in het RNAt.
Het is zeker dat er voor elk aminozuur minstens één RNAt is, maar het is niet even zeker of een enkel RNAt kan binden aan een enkel codon, of synoniemen onverschillig kan herkennen (vooral wanneer deze alleen voor de derde base verschillen).
We kunnen concluderen dat er gemiddeld drie synonieme codons zijn voor elk aminozuur, terwijl anticodons er minstens één en niet meer dan drie zijn.
Eraan herinnerend dat genen bedoeld zijn als enkelvoudige stukken van zeer lange polynucleotidesequenties van DNA, is het duidelijk dat het begin en het einde van het enkelvoudige gen noodzakelijkerwijs in het geheugen aanwezig moeten zijn.
BIOSYNTHESE VAN EIWITTEN
In verschillende delen van het DNA is er de opening van de dubbele keten en de synthese van de verschillende soorten RNA.
Tijdens de laadstap bindt het RNAt aan de aminozuren (eerder geactiveerd door het ATP en door het specifieke enzym). De biosynthetische "machine" is niet in staat om onjuist geladen tRNA's te "corrigeren".
Het RNAr splitst zich vervolgens in de twee subeenheden en geeft door binding aan de ribosomale eiwitten aanleiding tot de assemblage van de ribosomen.
Het RNAm, dat door het cytoplasma gaat, bindt zich aan de ribosomen en vormt zo het polysoom.Elk ribosoom, dat op de boodschapper stroomt, herbergt geleidelijk het RNAt dat complementair is aan de relatieve codons, waarbij de aminozuren worden genomen en ze aan de polypeptideketen in formatie binden.
Het relatief stabiele RNAt komt weer in de bloedsomloop. De ribosomen worden ook opnieuw gebruikt, waardoor het reeds samengestelde polypeptide vrijkomt.
De boodschapper, minder stabiel omdat het allemaal monocatenair is, wordt (door de ribonuclease) gesplitst in de samenstellende ribonucleotiden.
De cyclus gaat dus verder, waarbij de ene na de andere de polypeptiden op de boodschapper-RNA's worden gesynthetiseerd die door de transcriptie worden geleverd.